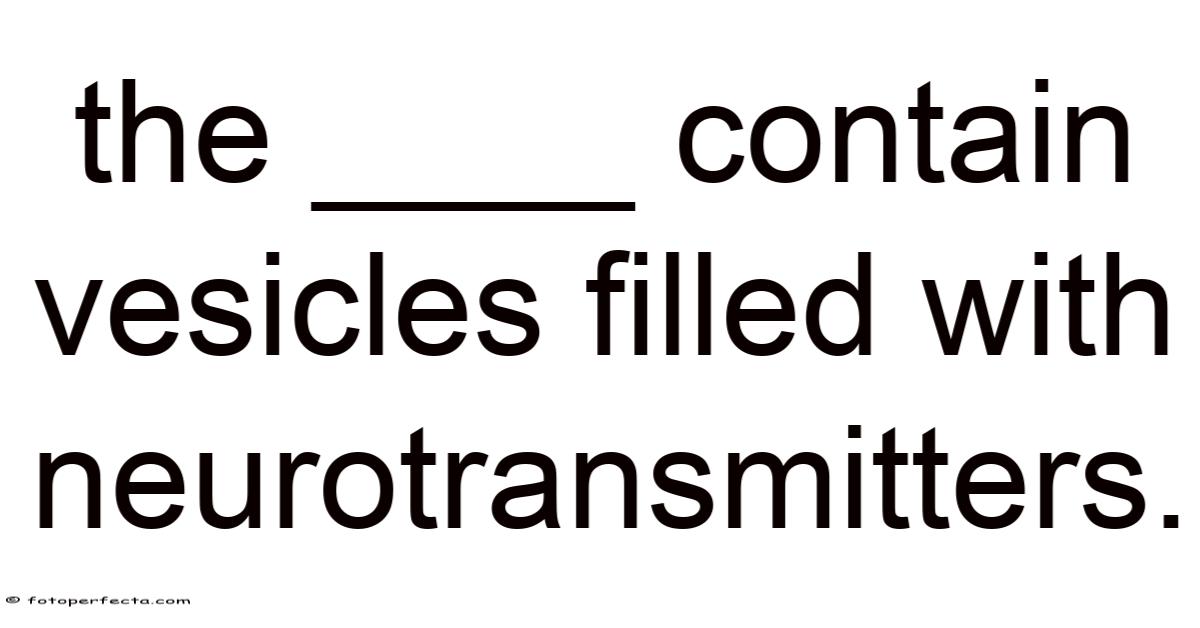
Introduction
Neurons communicate through a sophisticated chemical language, and the vesicles filled with neurotransmitters are the tiny messengers that make this dialogue possible. So these membrane‑bound packets are stored in a specialized region of the neuron known as the presynaptic terminal (or synaptic bouton). In real terms, when an electrical impulse—an action potential—reaches this terminal, the vesicles fuse with the plasma membrane and release their cargo into the synaptic cleft, where the neurotransmitters bind to receptors on the postsynaptic cell. Understanding how these vesicles are formed, loaded, transported, and released is essential for grasping everything from basic reflexes to complex behaviors such as learning, memory, and emotion regulation.
In this article we will explore the anatomy of the presynaptic terminal, the life cycle of neurotransmitter‑filled vesicles, the molecular machinery that drives exocytosis, and the clinical relevance of vesicle dysfunction. By the end, you will have a clear picture of why these microscopic containers are central to brain function and how they are being targeted in modern therapeutics.
Anatomy of the Presynaptic Terminal
Structure and Location
- Axon terminal (synaptic bouton): The distal swelling at the end of an axon, typically 0.5–2 µm in diameter.
- Active zone: A specialized area of the plasma membrane where vesicle docking and fusion occur. It contains a dense network of proteins that orchestrate release.
- Synaptic vesicles: Spherical, ~40 nm vesicles that store neurotransmitters. A single bouton can hold hundreds to thousands of vesicles, organized into distinct pools.
- Mitochondria and cytoskeleton: Provide ATP and structural support for vesicle trafficking.
Vesicle Pools
- Readily Releasable Pool (RRP): Vesicles already docked at the active zone, primed for immediate release upon calcium influx.
- Recycling Pool: Vesicles that can be quickly mobilized during moderate activity.
- Reserve Pool: Larger vesicles stored away from the active zone, recruited during intense or prolonged stimulation.
The balance among these pools determines a synapse’s firing reliability and plasticity.
Biogenesis of Neurotransmitter‑Filled Vesicles
1. Vesicle Formation in the Golgi Apparatus
Neurotransmitter vesicles originate from the trans‑Golgi network (TGN). Here, membrane proteins such as synaptophysin, synaptobrevin (VAMP), and vesicular transporters are incorporated into budding vesicles No workaround needed..
2. Loading of Neurotransmitters
Different neurotransmitters require distinct transport mechanisms:
- Amino acids (e.g., glutamate, GABA): Loaded via vesicular glutamate transporters (VGLUT1‑3) or vesicular GABA transporter (VGAT) using a proton gradient generated by the V‑ATPase pump.
- Monoamines (e.g., dopamine, serotonin, norepinephrine): Packaged by vesicular monoamine transporters (VMAT1‑2), also driven by the proton gradient.
- Peptides (e.g., substance P, oxytocin): Synthesized as larger precursors in the endoplasmic reticulum, then cleaved and packaged in dense‑core vesicles.
The proton electrochemical gradient (ΔpH and Δψ) is the key energy source, making the V‑ATPase indispensable for efficient loading Most people skip this — try not to..
3. Transport to the Axon Terminal
After formation, vesicles travel along microtubules powered by kinesin motor proteins. The journey can span several centimeters in motor neurons, yet vesicles maintain integrity thanks to a protective protein coat and continuous ATP supply.
The Release Process: From Calcium Influx to Exocytosis
Step‑by‑Step Overview
- Action Potential Arrival: Depolarization opens voltage‑gated N‑type and P/Q‑type calcium channels at the active zone.
- Calcium Influx: Intracellular Ca²⁺ concentration spikes from ~100 nM to >10 µM within milliseconds.
- Sensor Activation: The protein synaptotagmin binds Ca²⁺, undergoing a conformational change that triggers vesicle fusion.
- SNARE Complex Formation: Synaptobrevin (on the vesicle) pairs with syntaxin and SNAP‑25 (on the plasma membrane) to form a tight SNARE complex, pulling the membranes together.
- Membrane Fusion: The vesicle merges with the presynaptic membrane, creating a transient fusion pore.
- Neurotransmitter Release: Molecules diffuse across the synaptic cleft (≈20 nm wide) and bind to postsynaptic receptors.
- Vesicle Recycling: The membrane is retrieved via clathrin‑mediated endocytosis, reforming vesicles for another round.
Modulatory Factors
- Calcium Buffering: Proteins like calbindin modulate the effective Ca²⁺ concentration, shaping release probability.
- Phosphorylation: Kinases (e.g., PKA, PKC) phosphorylate SNARE components, adjusting vesicle priming and fusion rates.
- Synaptic Plasticity: Long‑term potentiation (LTP) and depression (LTD) alter vesicle pool sizes and release probability, underlying learning and memory.
Scientific Explanation of Vesicle Dynamics
Thermodynamics of Loading
The V‑ATPase pumps protons into the vesicle lumen, establishing a ΔpH of ~1.5–2 units and a membrane potential (Δψ) of ~30–60 mV. Neurotransmitter transporters exploit this gradient:
- Electrochemical coupling: For positively charged monoamines, the transporter uses the Δψ component; for negatively charged glutamate, the ΔpH component dominates.
- Kinetic advantage: The steep gradient ensures rapid loading, allowing the neuron to sustain high‑frequency firing without depletion.
Kinetic Modeling
Mathematical models describe vesicle release as a Markov process, with states representing docked, primed, and fused vesicles. In practice, the transition rates (k₁, k₂, k₃) are functions of calcium concentration and SNARE complex stability. Such models predict phenomena like paired‑pulse facilitation and depression, which are observable in electrophysiological recordings.
No fluff here — just what actually works.
Clinical Relevance
Neurological Disorders Linked to Vesicle Dysfunction
| Disorder | Vesicle‑Related Abnormality | Example of Therapeutic Target |
|---|---|---|
| Parkinson’s disease | Impaired VMAT2 activity → reduced dopamine storage | L‑DOPA (increases cytosolic dopamine) and VMAT2 enhancers under investigation |
| Schizophrenia | Dysregulated synaptic vesicle release probability, especially glutamatergic | NMDA‑receptor modulators and SNARE‑targeting peptides |
| Epilepsy | Excessive vesicle release leading to hyperexcitation | Synaptic vesicle protein 2A (SV2A) ligands such as levetiracetam |
| Depression | Altered serotonin vesicle loading (VMAT2) | Selective serotonin reuptake inhibitors (SSRIs) indirectly affect vesicle dynamics |
Worth pausing on this one.
Pharmacological Manipulation
- Botulinum toxins (BoNT): Cleave SNARE proteins, preventing vesicle fusion; used therapeutically for muscle spasticity.
- Tetanus toxin: Also targets SNAREs, leading to uncontrolled release; serves as a research tool.
- SV2A ligands: Bind to the vesicle protein SV2A, modulating release probability; basis for several antiepileptic drugs.
Understanding vesicle biology enables the design of drugs that fine‑tune neurotransmission without globally suppressing neuronal activity, offering more precise therapeutic windows.
Frequently Asked Questions
Q1. How many neurotransmitter molecules are inside a single vesicle?
A single synaptic vesicle typically contains ~4,000–5,000 molecules of a small‑molecule neurotransmitter (e.g., glutamate) or ~100–200 peptide molecules, reflecting differences in size and packing density.
Q2. Do all neurons use the same vesicle proteins?
While core SNARE components (synaptobrevin, syntaxin, SNAP‑25) are ubiquitous, accessory proteins such as Munc13, RIM, and Complexin vary among neuronal subtypes, contributing to distinct release kinetics.
Q3. Can vesicles be visualized directly?
Yes. Advanced imaging techniques—electron microscopy, super‑resolution fluorescence microscopy (STED, PALM), and live‑cell total internal reflection fluorescence (TIRF)—allow visualization of vesicle pools, docking, and fusion events with nanometer precision Worth keeping that in mind. No workaround needed..
Q4. What happens to vesicle membranes after exocytosis?
Membrane components are rapidly retrieved via endocytosis, predominantly through clathrin‑coated pits. This recycling conserves lipid and protein resources and maintains synaptic integrity.
Q5. Are there differences between excitatory and inhibitory vesicles?
Excitatory (glutamatergic) vesicles primarily contain VGLUT transporters, while inhibitory (GABAergic) vesicles use VGAT. Their release probabilities, docking proteins, and calcium sensitivities also differ, shaping the balance of excitation and inhibition in neural circuits.
Conclusion
The presynaptic terminal’s vesicles filled with neurotransmitters are far more than passive containers; they are dynamic, highly regulated machines that translate electrical signals into the chemical language of the brain. From their biogenesis in the Golgi apparatus to the exquisite calcium‑triggered fusion at the active zone, each step is orchestrated by a suite of proteins and gradients that ensure rapid, reliable communication.
Disruptions in any part of this vesicle cycle can lead to profound neurological and psychiatric conditions, underscoring the therapeutic potential of targeting vesicle‑related proteins. As imaging and molecular tools continue to evolve, our understanding of vesicle dynamics will deepen, opening new avenues for treating disorders rooted in synaptic dysfunction.
By appreciating the layered choreography of neurotransmitter‑filled vesicles, we gain insight into the very foundation of cognition, emotion, and behavior—a testament to how microscopic structures shape the grand tapestry of human experience.